Першоджерело: Follow-up NextQuestion
 Джерело зображення: Створено Unbounded AI
Джерело зображення: Створено Unbounded AI
Коли ми бачимо два набори лінійних решіток поруч один з одним, трохи зміщених один до одного, здається, що в нашій свідомості з’являється лінія, якої насправді не існує. Здається, що ця лінія згинається і простягається між двома наборами фактичних ліній (рисунок 1С).
 Рисунок 1: Варіант растрової галюцинації в шаховому порядку. А: Вазарелі (1970), Б: Зебре Вазарелі (1939), В: Класична ілюзія решітки в шаховому порядку. Джерело: Vasarely*
Рисунок 1: Варіант растрової галюцинації в шаховому порядку. А: Вазарелі (1970), Б: Зебре Вазарелі (1939), В: Класична ілюзія решітки в шаховому порядку. Джерело: Vasarely*
Таке просте розташування може створити дивовижний візуальний ефект, який ми називаємо ілюзією примикаючої решітки. Що ще цікавіше, так це те, що зі збільшенням кількості цих ліній наше сприйняття галюцинацій також стає сильнішим.
Також є дві ілюзії неонової дифузії, які також цікаві:
Ілюзія Еренштейна та ілюзія Валліна. При галюцинації Еренштейна набір радіальних ліній тягнеться назовні. Зовнішній напрямок лінії темний, а частина, що збирається до центру, світла. Якщо сфокусувати погляд на центрі лінії, то світла частина інтер’єру виглядає так, ніби вона розтікається в навколишню темну зону, утворюючи напівпрозоре коло.
 Малюнок 2: Галюцинація Еренштейна, де прямолінійний візерунок решітки, який зупиняється в певній точці, здається яскравішим у центрі, ніж на задньому плані, створюючи ілюзію кола. Джерело: Вікіпедія
Малюнок 2: Галюцинація Еренштейна, де прямолінійний візерунок решітки, який зупиняється в певній точці, здається яскравішим у центрі, ніж на задньому плані, створюючи ілюзію кола. Джерело: Вікіпедія
Ілюзія Валліна складалася з кіл по чотирьох кутах, одна частина кожного кола була світлішого кольору, а інші — чорного. Коли ми дивимося на ці кола, здається, що світлі частини розсіюються до центру, в кінцевому підсумку утворюючи напівпрозорий квадрат.
 Малюнок 3: Галюцинація Валіна, коли більш яскрава або темна частина центру поширюється до центру, в кінцевому підсумку утворюючи напівпрозорий квадрат. Джерело: Perception
Малюнок 3: Галюцинація Валіна, коли більш яскрава або темна частина центру поширюється до центру, в кінцевому підсумку утворюючи напівпрозорий квадрат. Джерело: Perception
Алгоритм сприйняття віртуального контуру
Вчені запропонували безліч теорій для пояснення алгоритмічного механізму віртуального сприйняття контурів, що запускається цими галюцинаціями.
Деякі школи вважають, що саме бічне придушення кольору підсилює контраст і змушує людей сприймати віртуальні лінії. Наприклад, коли нейрон збуджується яскравістю, він пригнічує нейрони в темніших областях навколо нього, через що яскраві ділянки здаються яскравішими, а темні – темнішими. Мозок може неправильно витлумачити посилення контрасту як чіткий контур і лінію[1] [2]。 Інша теорія припускає, що це може бути пов’язано з вищими функціями мозку, такими як доповнення мозку через знання та пам’ять. Коли наш мозок порівнює зображення, які ми бачимо, з минулими уявленнями, він може відчувати, що певних елементів не вистачає, тому він автоматично «завершує» ці відсутні частини, формуючи сприйняття віртуальних контурів [3]。 Існує також багато теорій, наприклад, що струс нерва може поєднувати фрагментарні риси, щоб сформувати уявлення про особливості, яких немає [4]Кортикальний коефіцієнт збільшення дбайливо ставиться до центрального поля зору і таким чином перебільшує характеристики певних подразників, наприклад …
Латеральне гальмування :* консервативний сигнальний механізм зіставлення, який керує утворенням різноманітних тонких візерунків, таких як плями та смугасті межі, у спочатку майже однорідних тканинах під час розвитку більшості організмів (посилання 9)
Що стосується нейрофізіологічного механізму віртуального контурного сприйняття, то, незважаючи на те, що вчені провели безліч досліджень, до сих пір існує безліч суперечок. Одна з конкуруючих моделей припускає, що цей нейрофізіологічний механізм виникає в нижній зоровій корі (V1/V2), а потім впливає на вищу кору знизу вгору, тоді як інша припускає, що цей процес спочатку локалізується у вищій корі, насамперед у латеральній потиличній корі (LOC), а потім впливає на V1/V2 зверху вниз[5] 。
Що стосується ілюзії неонової колірної дифузії, деякі дослідження показують, що V3 і V4 мають значний вплив на явища заповнення кольору, пов’язані з формою[6] [7]; У той же час є деякі дослідження, які свідчать про те, що кортикальна активність, пов’язана з появою кольору, що виникає в результаті індукованого краєм заповнення або однорідної кольоровості поверхні, пов’язана з комплементарними патернами активності в дорсальному (V3A і V3B/KO) і вентральному зоровому потоці (V3v, hV4 і LO). (Малюнок 4).
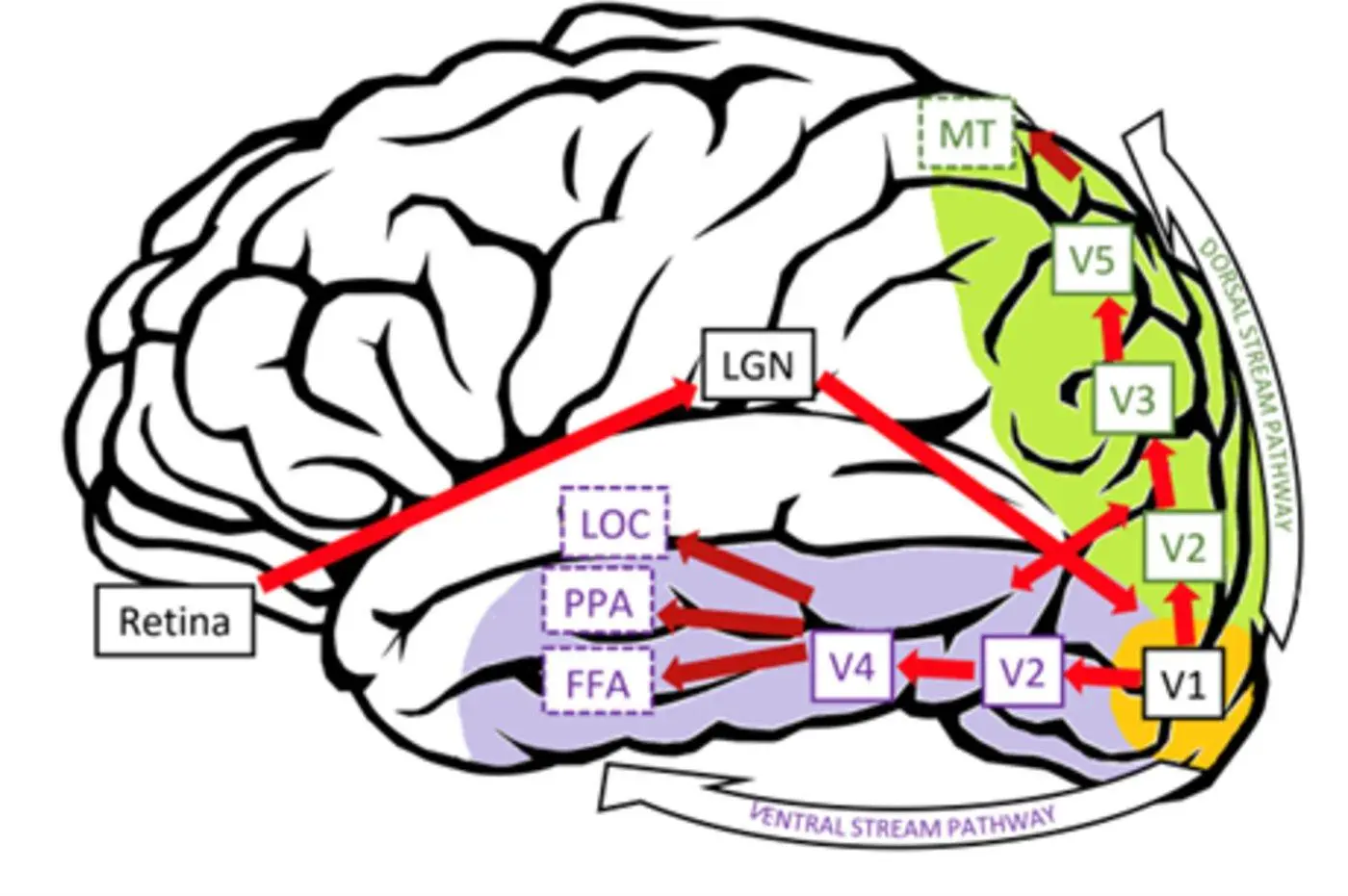 Рисунок 4: Різні функціональні області та шляхи в зоровій корі. Джерело: посилання 7.
Рисунок 4: Різні функціональні області та шляхи в зоровій корі. Джерело: посилання 7.
Хоча ці теорії дають деяке пояснення алгоритмічним і фізіологічним механізмам галюцинацій, все ще неясно, як ці зорові галюцинації представлені в свідомості, і здається неможливим їх перевірити.
Коли ми бачимо галюцинації, чи дійсно ми їх «бачимо»?
Чи можемо ми бути впевнені, що використовуємо «галюцинації» для характеристики стимулів, які викликають галюцинації?
Реконструкція галюцинаторних образів
Нещодавня стаття, опублікована в Science Advances, перевірила два типи зорових галюцинацій, згаданих вище, і успішно реконструювала зображення ліній і кольорів, що відповідають галюцинаторному досвіду, продемонструвавши ефективність моделі в матеріалізації суб’єктивних переживань[8] 。
 ▷Малюнок 1: Титульна сторінка статті. Джерело: Посилання 8
▷Малюнок 1: Титульна сторінка статті. Джерело: Посилання 8
В експерименті дослідники попросили 7 випробовуваних переглянути три або більше візуальних зображень галюцинацій і відповідні їм контрольні зображення і позитивні контрольні зображення. ** Для кожної ілюзії змінено контрольну картинку. Наприклад, контрольне зображення перемежованої растрової ілюзії сильно зменшує щільність огорожі, контрольне зображення галюцинації Еренштейна, в якому хвостовий кінець лінії, яка спочатку викликала галюцинацію, світліша, тоді як позитивне керуюче зображення змінює променистий центр лінії на світлове коло, а контрольне зображення галюцинації Валліна видаляє чорну частину галюцинаторного стимулу (початковий візуальний стимул), тоді як позитивне контрольне зображення змінює центральну частину на світліший кольоровий квадрат (докладніше див. Рисунок 5).
Зображення блимали з частотою 0,625 Гц по 8 секунд кожне і повторювалися по 20 разів. Протягом усього експерименту функціональна магнітно-резонансна томографія (фМРТ) використовувалася для реєстрації активності мозку, коли учасники дивилися зображення.
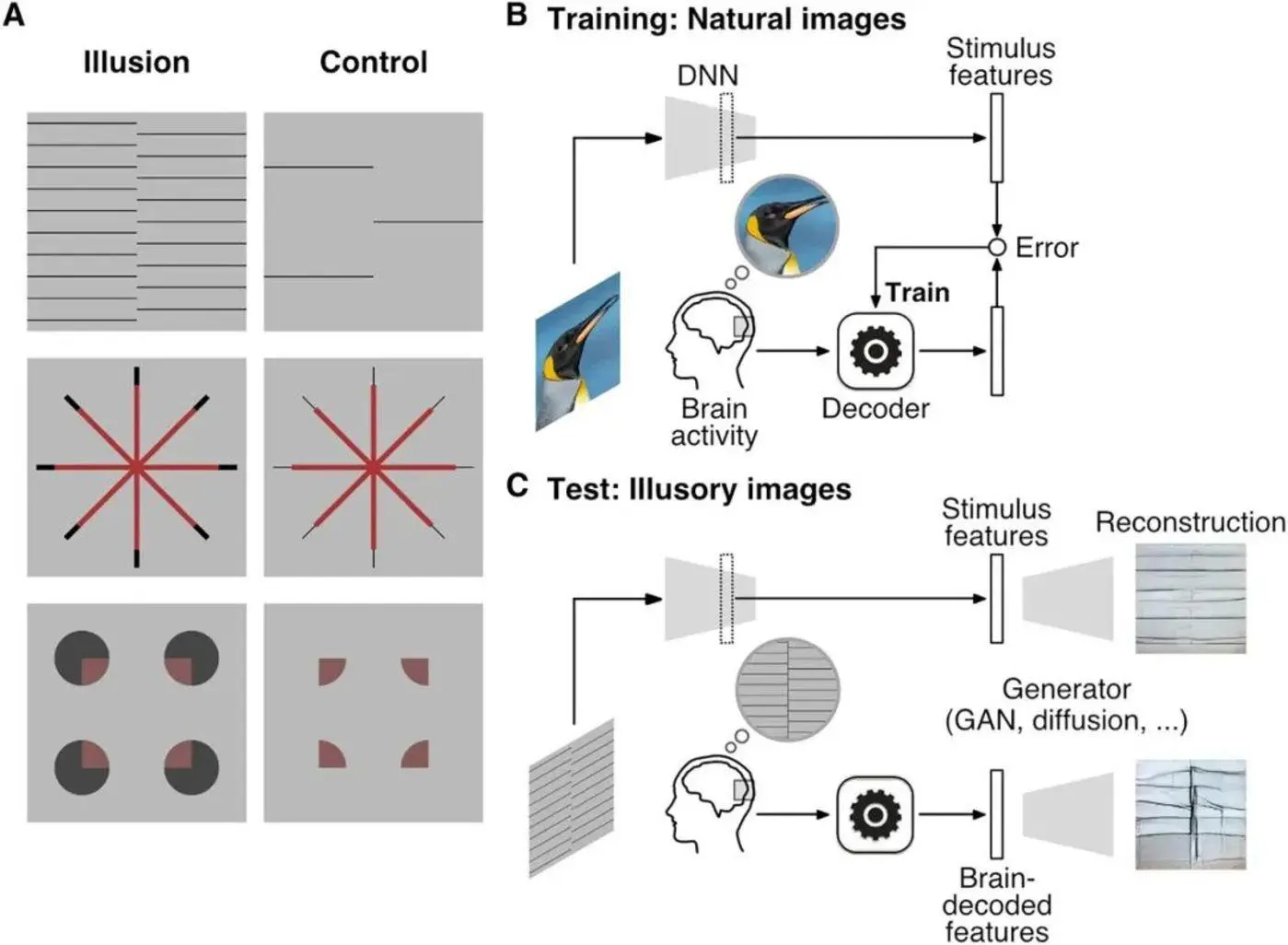 Малюнок 5: Ліва колонка малюнка А - це стимул, який викликає галюцинацію, а права колонка малюнка А - це керуючий стимул, що відповідає стимулу, який викликає галюцинацію. Перший рядок фігури А - це растрова галюцинація в шаховому порядку, другий рядок фігури А - галюцинація Еренштейна, а третій рядок фігури А - галюцинація Валліна. На малюнку В показаний процес тренування DNN за допомогою природних зображень. На малюнку C показана активність мозку, показана в даних декодування ДНК фМРТ як особливості стимулу, і процес, за допомогою якого генератор генерує картину на основі результатів декодування ДНН. Джерело: Наука
Малюнок 5: Ліва колонка малюнка А - це стимул, який викликає галюцинацію, а права колонка малюнка А - це керуючий стимул, що відповідає стимулу, який викликає галюцинацію. Перший рядок фігури А - це растрова галюцинація в шаховому порядку, другий рядок фігури А - галюцинація Еренштейна, а третій рядок фігури А - галюцинація Валліна. На малюнку В показаний процес тренування DNN за допомогою природних зображень. На малюнку C показана активність мозку, показана в даних декодування ДНК фМРТ як особливості стимулу, і процес, за допомогою якого генератор генерує картину на основі результатів декодування ДНН. Джерело: Наука
Далі дослідники фМРТ, зібрані дані мозку, були передані в навчену глибоку нейронну мережу (DNN) для розшифровки. Цей DNN є варіантом AlexNet, моделі BAIR/BVLC CaffeNet, розробленої Алексом Крижевським, директором Open AI. Це згорткова нейронна мережа, що використовує алгоритм прямого поширення, який був попередньо навчений класифікувати 1000 об’єктів у базі даних зображень ImageNet.
** Результати показали, що образи, реконструйовані з ментальних уявлень галюцинаторних стимулів, проявляють галюцинаторні властивості. На відміну від них, образи, реконструйовані безпосередньо з природи самого подразника, не виявляють галюцинаторних властивостей. Це також показує, що природа галюцинації, реконструйованої з ментальної репрезентації, побудована не природою і обробкою ДНМ, а природою самої ментальної репрезентації (див. Рис. 6).
 Малюнок 6: Крайній лівий стовпчик - це картина стимулу. Другий стовпець зліва - це зображення ДНК, реконструйоване з даних самого зображення стимулу в крайньому лівому стовпці. На малюнках 3-6 представлені уявні уявлення, реконструйовані з мозкової активності, що генерується крайнім лівим стимульним зображенням суб’єкта 1. На малюнках 7-10 представлені ментальні уявлення, реконструйовані з мозкової активності, що генерується крайніми лівими стимульними образами двох пар. Лінії 1-3 малюнка 1-3 є зображеннями ілюзії решітки в шаховому порядку. Перший ряд присвячений ілюзії решітки в шаховому порядку з двох колон решітчастих решіток, розташованих уздовж 90 градусів, другий ряд - ілюзії решітки в шаховому порядку з двох колон решітки, розташованих уздовж 45 градусів, а третій ряд - про управління ілюзією решітки в шаховому порядку. Рядки 4-7 – це зображення галюцинацій Еренштейна. Четвертий і шостий рядки присвячені галюцинації Еренштейна, чотири мають 8 струн, а шість - 4 рядки. Лінії 5 і 7 присвячені управлінню галюцинацією Еренштейна, п’ять мають 8 ліній, а сім — 4 лінії. Рядки 8-9 малюнка 8-9 є зображеннями галюцинацій Валліна. Восьмий акт - галюцинація Валліна, а дев’ятий акт - контролюючий. Джерело: Наука
Малюнок 6: Крайній лівий стовпчик - це картина стимулу. Другий стовпець зліва - це зображення ДНК, реконструйоване з даних самого зображення стимулу в крайньому лівому стовпці. На малюнках 3-6 представлені уявні уявлення, реконструйовані з мозкової активності, що генерується крайнім лівим стимульним зображенням суб’єкта 1. На малюнках 7-10 представлені ментальні уявлення, реконструйовані з мозкової активності, що генерується крайніми лівими стимульними образами двох пар. Лінії 1-3 малюнка 1-3 є зображеннями ілюзії решітки в шаховому порядку. Перший ряд присвячений ілюзії решітки в шаховому порядку з двох колон решітчастих решіток, розташованих уздовж 90 градусів, другий ряд - ілюзії решітки в шаховому порядку з двох колон решітки, розташованих уздовж 45 градусів, а третій ряд - про управління ілюзією решітки в шаховому порядку. Рядки 4-7 – це зображення галюцинацій Еренштейна. Четвертий і шостий рядки присвячені галюцинації Еренштейна, чотири мають 8 струн, а шість - 4 рядки. Лінії 5 і 7 присвячені управлінню галюцинацією Еренштейна, п’ять мають 8 ліній, а сім — 4 лінії. Рядки 8-9 малюнка 8-9 є зображеннями галюцинацій Валліна. Восьмий акт - галюцинація Валліна, а дев’ятий акт - контролюючий. Джерело: Наука
** Дослідження унікальне тим, що зображення розшифровуються з даних з усієї зорової кори, записаних на фМРТ, і що реконструйовані ментальні репрезентації, як правило, демонструють риси, які мають більш галюцинаторний характер, ніж сам стимул. **
У разі переплетених растрових галюцинацій ілюзорна лінія в галюцинаторному стимулі виявляється набагато більш вираженою, ніж лінії, що покриваються будь-яким подразником як таким, в той час як психічні уявлення, реконструйовані з контрольного стимулу, не вказують на це; у разі неонових колірних дифузійних галюцинацій у галюцинації Еренштейна частка дифузії кольору значно більша, ніж у контрольній частині; у галюцинації Валліна, хоча контрольний стимул все ще показує обриси дифузійної форми, частина дифузії кольору все ж значно більша, ніж у контрольній частині. Більш того, управління самим стимулом призначене тільки для того, щоб пригнічувати дифузію кольору, а не контур форми, тому в разі галюцинації Уолліна галюцинаторна властивість сильніше, ніж стимул в уявному уявленні, і ця властивість все ще дійсна.
Кількісна оцінка ілюзії реконструкції
Інтенсивність галюцинацій не тільки відбивається на реконструйованих образах, але і може бути кількісно оцінена за допомогою цілого ряду методів. У даній роботі представлений детальний кількісний аналіз інтенсивності галюцинацій в психічних уявленнях, викликаних галюцинаціями в шаховому порядку і галюцинаторними стимулами неонової дифузії.
Дослідники використовували радонове перетворення для аналізу галюцинаторних властивостей ілюзії решітки в шаховому порядку, тобто інтенсивності віртуальних ліній між ґратами. ** Як широко використовуваний метод аналізу рентгенографічних даних, перетворення Ладунга може математично реконструювати форму вихідного об’єкта за інтегральними значеннями (наприклад, щільністю, масою тощо), отриманими шляхом проектування об’єкта з усіх напрямків, тоді як можна знати лише опис сторони. **
У цьому дослідженні перетворення Ладунга застосовується до аналізу зображення, а основний напрямок розподілу ліній у реконструйованому графі виходить шляхом підсумовування «значень пікселів», отриманих при проектуванні зображення з усіх напрямків. Експерименти показали, що при всіх даних про зорову кору на фМРТ переважні напрямки на реконструйованих зображеннях показують бімодальний розподіл, при якому близько 61,1% переважаючих напрямків ближче до переважного напрямку галюцинації, ніж до візуально індукованого.
З графіка видно, що вертикальні абсолютні значення в бімодальному розподілі в головному напрямку реконструйованого зображення вищі і більш кластеризовані, ніж величини в поперечних абсолютних значеннях. У міру зменшення числа решіток (ліній) в галюцинаторних подразниках вертикальна абсолютна величина починає зменшуватися щодо поперечного абсолютного значення в межах бімодального розподілу основного напрямку в реконструйованому уявленні подразника уявним уявленням, причому поперечне абсолютне значення значно перевищує вертикальне абсолютне значення в контрольному стимулі. ** Цей результат підтверджує теорію про те, що зменшення числа решіток (ліній) зменшить ступінь галюцинацій. **
При цьому основний напрямок зони галюцинаторної концентрації близький до основного напрямку реконструйованого зображення, в той час як основний напрямок негалюцинаторной зони (області, де в основному зосереджена зорова індукція) не близький до основного напрямку реконструйованого образу. Це доводить, що область, в якій розташовані галюцинації в реконструйованому зображенні, знаходиться між двома наборами ґрат, як це сприймається (див. рис. 7B, C, D, E).
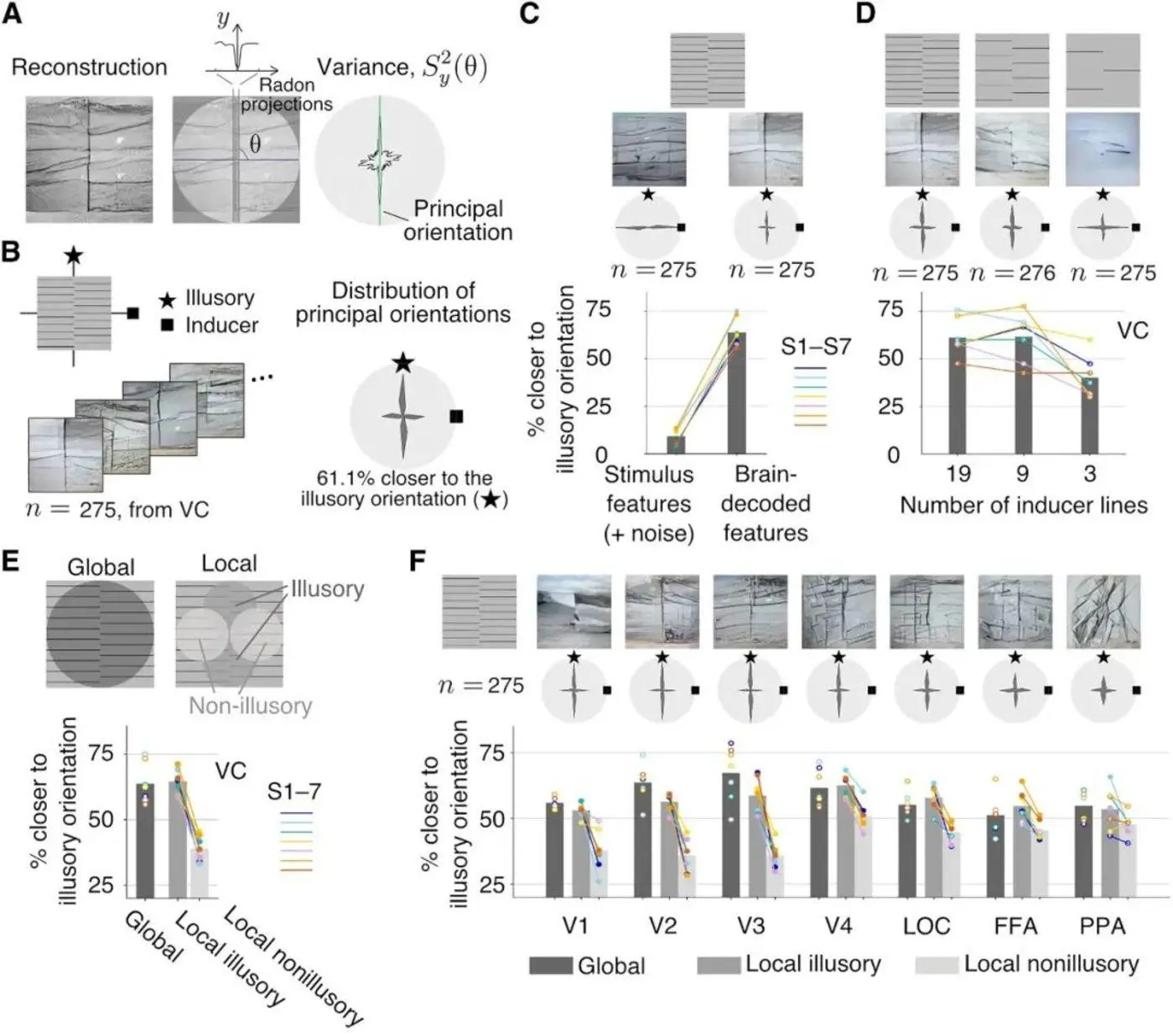 Малюнок 7: Малюнок А ілюструє процес перетворення Ладуна. На малюнку В видно, що 61,1% основних напрямків реконструйованої картини ближче до основного напрямку галюцинацій щодо індукованого основного напрямку. На малюнку С видно, що основний напрямок стимульної оцінки значно ближче до основного напрямку індукції. На малюнку D видно, що чим менше число ліній, тим ближче основний напрямок малюнка до основного напрямку спонукання. На малюнку E показано, що основний напрямок області галюцинаторної концентрації також близький до основного напрямку реконструйованого зображення На малюнку F показана ментальна репрезентація, реконструйована з даних, отриманих з різних областей зорової кори, а V1-V3 більш галюцинаторна. Джерело: Наука
Малюнок 7: Малюнок А ілюструє процес перетворення Ладуна. На малюнку В видно, що 61,1% основних напрямків реконструйованої картини ближче до основного напрямку галюцинацій щодо індукованого основного напрямку. На малюнку С видно, що основний напрямок стимульної оцінки значно ближче до основного напрямку індукції. На малюнку D видно, що чим менше число ліній, тим ближче основний напрямок малюнка до основного напрямку спонукання. На малюнку E показано, що основний напрямок області галюцинаторної концентрації також близький до основного напрямку реконструйованого зображення На малюнку F показана ментальна репрезентація, реконструйована з даних, отриманих з різних областей зорової кори, а V1-V3 більш галюцинаторна. Джерело: Наука
Дослідники також використовували лінійну регресію для аналізу галюцинаторної природи галюцинацій Еренштейна та Валліна, тобто ступеня, до якого колір дифундує назовні (утворюючи напівпрозору площину) від ліній, заданих стимулом. Вони побудували «карту почервоніння» на основі значень RGB пікселів у кожному зображенні, відповідно, для реконструйованих зображень галюцинаторних ментальних репрезентацій, контрольних ментальних репрезентацій та позитивних контрольних ментальних репрезентацій, а також для стимульних зображень та очікуваних суб’єктивних галюцинаторних образів.
Далі дослідники спробували підігнати карту почервоніння ментальної репрезентації з картою почервоніння стимулу та очікуваною суб’єктивною галюцинацією. Очікувані суб’єктивні галюцинації і стимули мають відповідний коефіцієнт (β1 і β2), і ці коефіцієнти представляють їх відповідну вагу в реконструйованих психічних репрезентаціях. Тобто, чим більше частин реконструюють червону частину ментального уявлення, тим більше коефіцієнт β1. Це пояснюється тим, що тільки суб’єктивні галюцинації, які є більш червоними в момент сприйняття, мають більшу частку суб’єктивних галюцинацій, які роблять уявні уявлення більш червоними (див. Рис. 8C, D, G, F).
Результати лінійної регресії показали, що виходячи з усіх даних про зорову кору з фМРТ, суб’єктивні галюцинаційні коефіцієнти обох стимулів, що викликали дифузні галюцинації неонового кольору, були більшими, ніж у контрольного стимулу, і меншими, ніж у позитивного контрольного стимулу. Однак суб’єктивний коефіцієнт галюцинації галюцинації Уоллінга, хоча і більше, ніж у контрольної, був значно менше, ніж у галюцинації Еренштейна (див. Рис. 8Е, Ф, І, Ж).
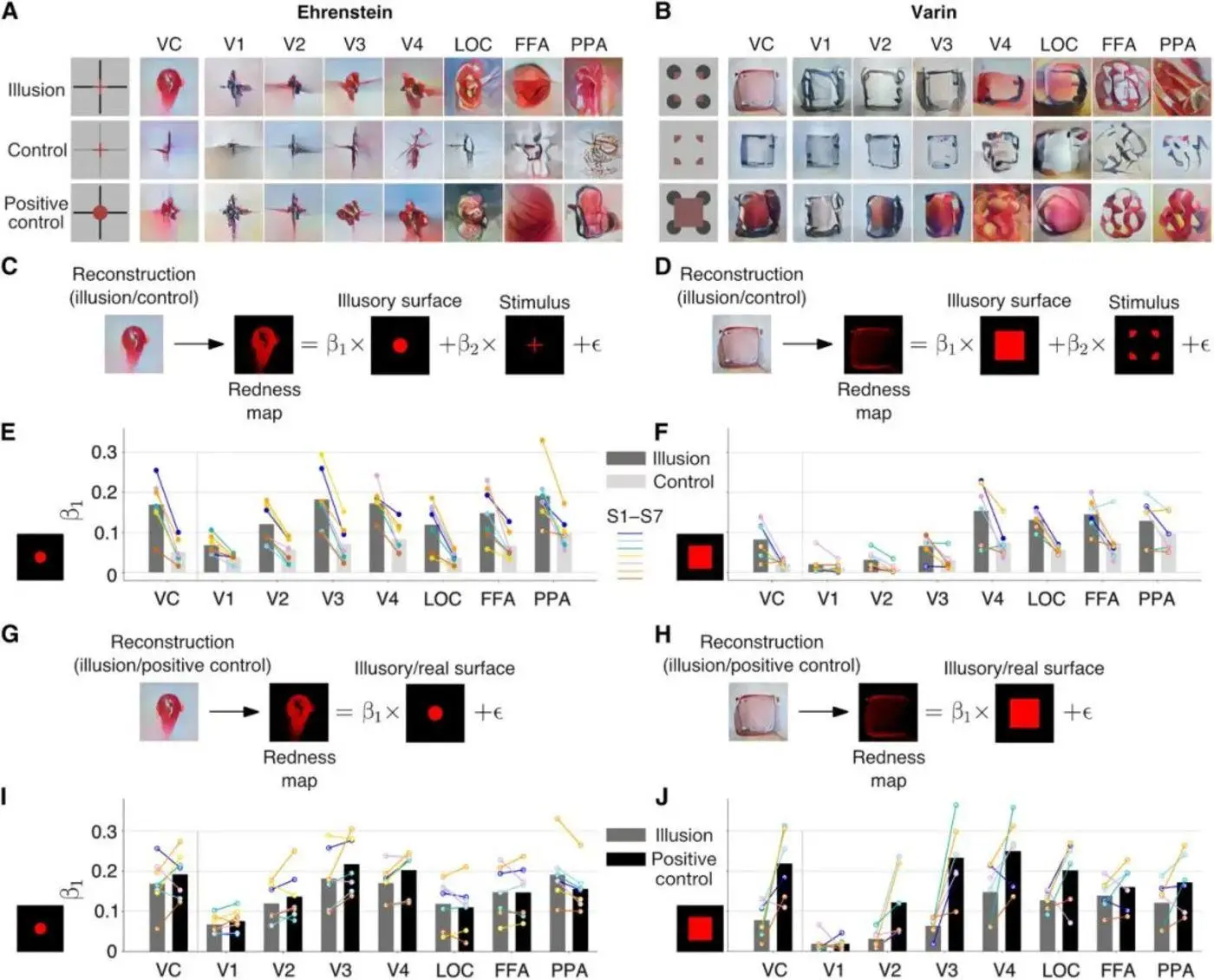 Малюнок 8: Малюнок А ілюструє галюцинацію Еренштейна, з її контролем і позитивним контролем в різних областях зорової кори. Панель B ілюструє галюцинацію Валліна, представлення її контролю та позитивного контролю в різних областях зорової кори. Панель C показує, як обчислюється лінійна регресія, порівнюючи галюцинації Еренштейна та графік почервоніння під їхнім контролем. На панелі D показано, як обчислюється лінійна регресія контрастних галюцинацій Валліна і карта почервоніння, яку вони контролюють. Панель Е показує частку галюцинаторних властивостей галюцинацій Еренштейна по відношенню до властивостей контрольних галюцинацій в кожній області зорової кори. На малюнку F показана частка галюцинаторних властивостей Валліна по відношенню до контрольних галюцинаторних властивостей в різних областях зорової кори. Панель G показує, як обчислюється лінійна регресія для порівняння галюциєнта Еренштейна з його позитивно контрольованою діаграмою почервоніння. На панелі H показано, як обчислюється лінійна регресія контрастних галюцинацій Валліна та їх позитивно контрольованих графіків почервоніння. На малюнку I показана частка галюцинаторних властивостей галюцинацій Еренштейна в кожній області зорової кори по відношенню до позитивних контрольних галюцинаторних властивостей. На малюнку J показана частка галюцинаторних властивостей Валліна в кожній області зорової кори по відношенню до позитивних контрольних галюцинаторних властивостей. Джерело: Наука
Малюнок 8: Малюнок А ілюструє галюцинацію Еренштейна, з її контролем і позитивним контролем в різних областях зорової кори. Панель B ілюструє галюцинацію Валліна, представлення її контролю та позитивного контролю в різних областях зорової кори. Панель C показує, як обчислюється лінійна регресія, порівнюючи галюцинації Еренштейна та графік почервоніння під їхнім контролем. На панелі D показано, як обчислюється лінійна регресія контрастних галюцинацій Валліна і карта почервоніння, яку вони контролюють. Панель Е показує частку галюцинаторних властивостей галюцинацій Еренштейна по відношенню до властивостей контрольних галюцинацій в кожній області зорової кори. На малюнку F показана частка галюцинаторних властивостей Валліна по відношенню до контрольних галюцинаторних властивостей в різних областях зорової кори. Панель G показує, як обчислюється лінійна регресія для порівняння галюциєнта Еренштейна з його позитивно контрольованою діаграмою почервоніння. На панелі H показано, як обчислюється лінійна регресія контрастних галюцинацій Валліна та їх позитивно контрольованих графіків почервоніння. На малюнку I показана частка галюцинаторних властивостей галюцинацій Еренштейна в кожній області зорової кори по відношенню до позитивних контрольних галюцинаторних властивостей. На малюнку J показана частка галюцинаторних властивостей Валліна в кожній області зорової кори по відношенню до позитивних контрольних галюцинаторних властивостей. Джерело: Наука
Зона зору, що відповідає галюцинації
Дослідники не тільки реконструювали ментальне представлення галюцинацій у всій зоровій корі, але й реконструювали психологічні репрезентації галюцинацій у кожній конкретній зоровій області кори.
Ці зорові області розташовані в порядку від середини та вниз за течією до верхніх меж зорового шляху (від первинного до вищого функціонування), включаючи первинну зорову кору (V1), вторинну зорову кору (V2), третинну зорову кору (V3), четвертинну зорову кору (V4), латеральний потиличний частковий комплекс (LOC), веретеноподібну звивину (FFA) та парагіппокампальну область (PPA).
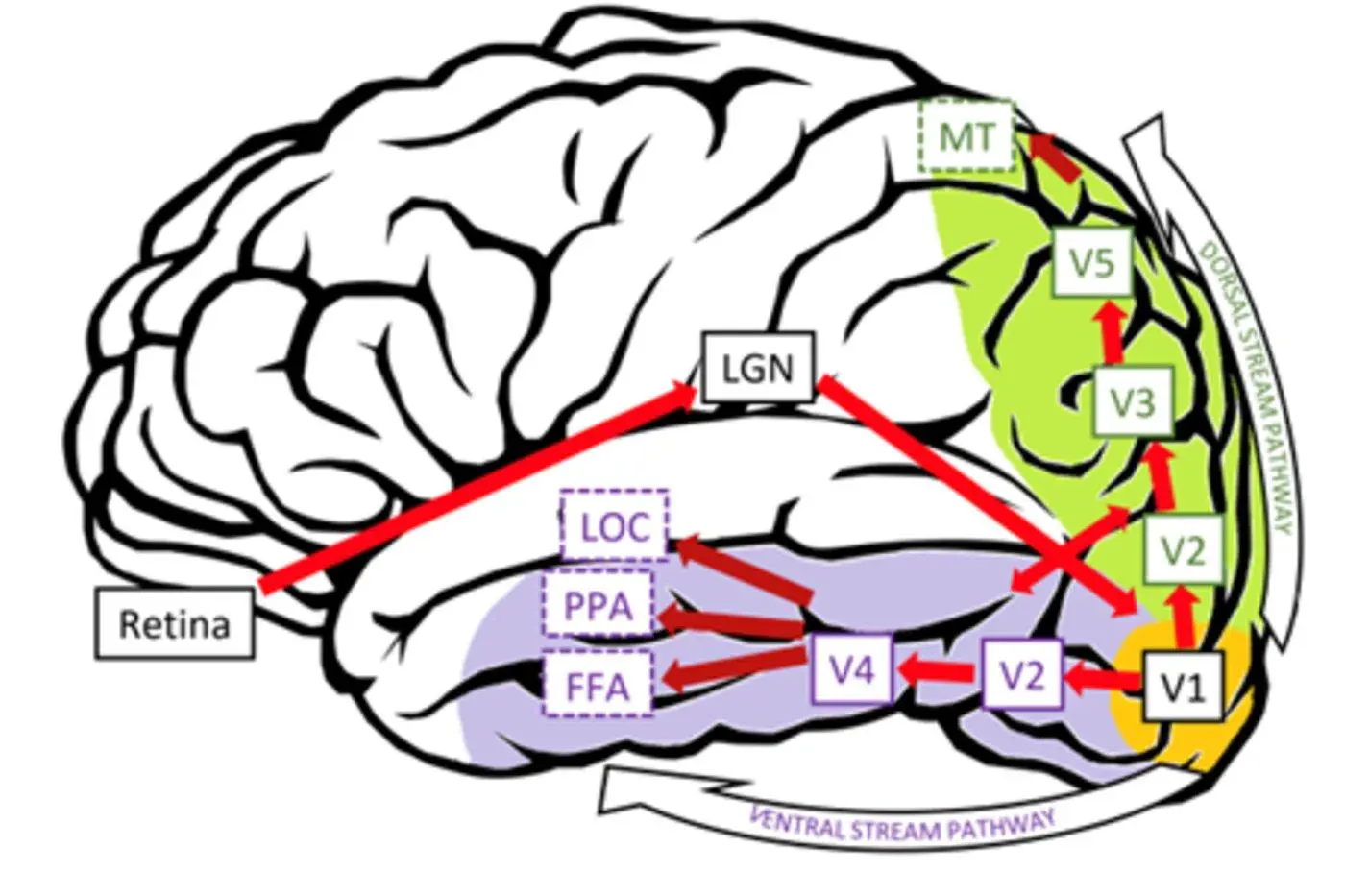 Малюнок 4: Діаграма різних функціональних областей і шляхів в зоровій корі. Джерело: посилання 7.
Малюнок 4: Діаграма різних функціональних областей і шляхів в зоровій корі. Джерело: посилання 7.
Після вилучення даних фМРТ з різних областей зорової кори та розшифровки та реконструкції ментальних репрезентацій відповідно, дослідники виявили, що для растрових галюцинацій, розташованих у шаховому порядку, галюцинаторні властивості в основному відображалися в V1-V3 лише на зображеннях, а галюцинаторні властивості та стимули в областях V4 і вище не були чіткими. Кількісно оцінивши основні напрямки реконструкції зображення в кожній області, дослідники виявили, що галюцинаторна природа досягає піку між V2 і V4 у всій області зображень і досягає піку між V1 і V3 в області галюцинаторної концентрації. Ці результати свідчать про те, що характеристика ліній в основному зосереджена на ранній зоровій області (рис. 7F).
Маючи справу з галюцинаціями, що розповсюджують неонові кольори, зорова кора використовує різні стратегії. Дослідження показало, що галюцинаторний характер галюцинацій Еренштейна значно відображався в кожній області зорової кори, що узгоджувалося з представленням неонового кольору позитивно контролюючого стимулу в кожній області зорової кори, в той час як галюцинаторний характер галюцинацій Уоллінга лише значно відображався в області зорової кори на V4 і вище, в той час як неоновий колір позитивно контролюючого стимулу також був значно відображений в V2 і V3. У поєднанні з результатами того, що коефіцієнт суб’єктивної галюцинації галюцинації Валліна був меншим, ніж коефіцієнт суб’єктивної галюцинації галюцинації Еренштейна, це вказує на те, що навіть якщо природа галюцинації однакова, різні налаштування індукції викличуть різні методи обробки інформації в зоровій корі. **
Коли у нас є зорові галюцинації, це вплив зверху вниз первинної зорової кори з вищих областей вниз для побудови галюцинацій, чи це викликано тим, що первинна зорова кора знизу вгору впливає на вищі області? Хоча ця стаття представляє лише уявлення, передбачені в попередніх теоріях, з іншої точки зору, замість того, щоб намагатися побудувати причинно-наслідковий зв’язок між мозковою активністю та природою галюцинацій, її яскрава презентація все ще дає нам деякі підказки щодо того, як суб’єктивний досвід активний у мозку.
Очевидно, що ми не можемо просто використовувати одну область мозку для реконструкції всього галюцинаторного досвіду, і всі індуковані сигнали будуть впливати на механізм одного і того ж галюцинаторного досвіду. Здається, що потрібна більш повна теоретична база, щоб зрозуміти складний механізм виникнення галюцинацій.
Наступного разу, коли ви побачите ці сліпучі галюцинаторні образи, ви можете подумати про галюцинації, створені цими моделями, про чудове змагання «просунутий проти початківця», яке може розігруватися у вашому мозку, а потім посміхнутися і здивуватися диву пізнання.
Посилання:
[1] . Елліотт, С. Л., і Шевелл, С. К. (2013). «Сприйнята сегментація центру від оточення лише ілюзорними контурами викликає хроматичне латеральне гальмування». Це дослідження припускає, що ілюзорні контури можуть викликати просторовий антагонізм «центр-оточення», щоб змінити зовнішній вигляд кольору, вказуючи на кортикальний нейронний локус для латерального гальмування
[2] . Пак А, Рю Е, Лі С, Чубикін А.А. Зворотний зв’язок зверху вниз контролює кортикальне представлення ілюзорних контурів у первинній зоровій корі миші. Дж. 2020 Січ 15; 40(3):648-660. doi: 10.1523/JNEUROSCI.1998-19.2019. Електронна публікація 2 грудня 2019 р. PMID: 31792152; PMCID: PMC6961994.
[3] . Мюррей М.М., Германн С. Ілюзорні контури: вікно в нейрофізіологію конструювання сприйняття. Тенденції Cogn Sci. 2013 Вересень; 17(9):471-81. DOI: 10.1016/J.tics.2013.07.004. Електронна публікація 6 серпня 2013 р. PMID: 23928336.
[4] . Гоув, А., Гроссберг, С., і Мінголла, Е. (1995). Сприйняття яскравості, ілюзорні контури та кортикогенікуляційний зворотний зв’язок Visual Neuroscience, 12(6), 1027-1052. DOI:10.1017/S0952523800006702
[5] . Ф. Кнебель, М. М. Мюррей, На шляху до вирішення суперечливих моделей ілюзорної обробки контурів у людини. Neuroimage 59, 2808–2817 (2012).
[6] . С. В. Хонг, Ф. Тонг, Нейронна репрезентація контингентного заповнення кольорів у ранній зоровій корі. 17, 10 (2017).
[7] . П. Жерардін, К. Аббатекола, Ф. Девінк, Х. Кеннеді, М. Дохат, К. Кноблаух, Нейронні ланцюги для заповнення кольорів на великій відстані. Нейрообраз 181, 30–43 (2018).
[8] . Fan L. Cheng et al., Реконструкція зорових ілюзорних переживань з активності людського мозку.Sci. Adv.9,eadj3906(2023). DOI:10.1126/sciadv.adj3906
[9] Коен, М. та ін (2010) «Динамічні філоподії передають переривчасту сигналізацію дельта-виїмки для уточнення патерну під час латерального гальмування», Клітина розвитку, 19(1), с. 78–89. DOI:10.1016/j.devcel.2010.06.006

